Introduction of AAV
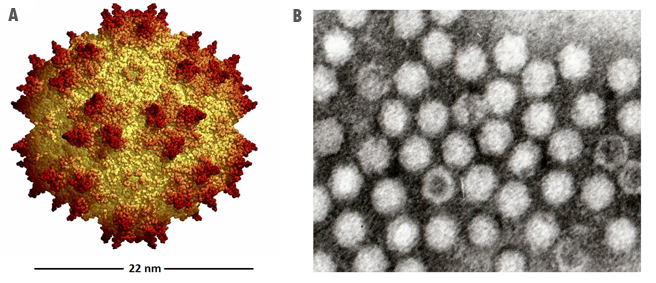
Adeno-associated virus (AAV) is a small virus which infects humans and some
other primate species. AAV belongs to the genus Dependovirus, which in turn
belongs to the family Parvoviridae. The virus is a small (20 nm)
replication-defective, nonenveloped virus. AAV is not currently known to
cause disease and consequently the virus causes a very mild immune response.
Wild type AAV
can infect both dividing and non-dividing cells and may incorporate its genome
into that of the host cell. These features make AAV a very attractive candidate
for creating viral vectors for gene therapy, and for the creation of isogenic
human disease models.
|
|
Serotype |
CNS/Retina |
Heart |
Lung |
Liver |
Skeletal Muscle |
|
AAV1 |
X |
X |
X |
|
X |
|
AAV2 |
X |
|
|
X |
X |
|
AAV3 |
X |
X |
|
X |
|
|
AAV4 |
X |
X |
|
|
|
|
AAV5 |
X |
|
X |
|
|
|
AAV6 |
|
X |
X |
X |
X |
|
AAV7 |
X |
|
|
X |
X |
|
AAV8 |
X |
|
|
X |
X |
|
AAV9 |
X |
X |
X |
X |
X |
|
AAV10 |
X |
|
X |
|
|
AAV immunology
AAV is of particular interest to gene
therapists due to its apparent limited capacity to induce immune responses in
humans, a factor which should positively influence vector transduction
efficiency while reducing the risk of any immune-associated pathology. The innate immune response to the AAV vectors has been characterised in
animal models. Intravenous administration in mice causes transient production of
pro-inflammatory cytokines and some infiltration of neutrophils and other
leukocytes into the liver.
- Cell-mediated
The cell-mediated response to the virus and to vectors is poorly
characterised and has been largely ignored in the literature as recently as
2005. Clinical trials using an AAV2-based vector to treat haemophilia B seem to
indicate that targeted destruction of transduced cells may be occurring.
Combined with data that shows that CD8+ T-cells can recognise elements of the
AAV capsid in vitro, it appears that there may be a cytotoxic T lymphocyte
response to AAV vectors. Cytotoxic responses would imply the involvement of CD4+
T helper cells in the response to AAV and in vitro data from human studies
suggests that the virus may indeed induce such responses including both Th1 and
Th2 memory responses. A number of candidate T cell stimulating epitopes have
been identified within the AAV capsid protein VP1, which may be attractive
targets for modification of the capsid if the virus is to be used as a vector
for gene therapy.
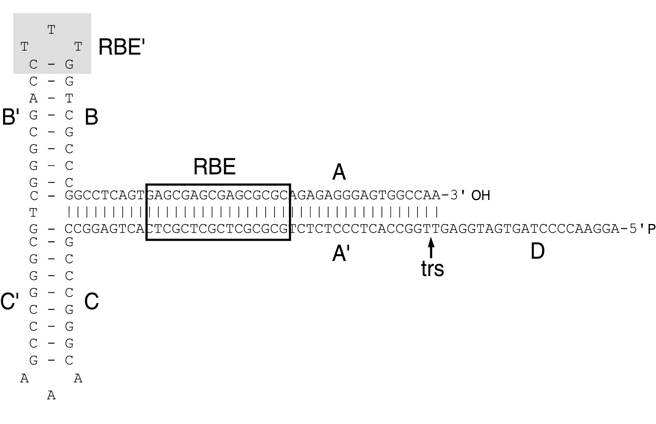
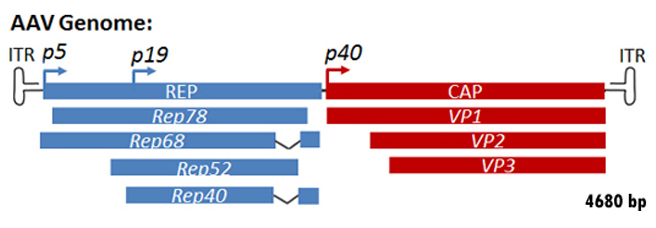
How Is The Recombinant AAV (rAAV) Packaged?
Unlike wild type AAV, the recombinant AAV (rAAV) fails to integrate its genome
to host cells. With the newly developed rAAV packaging technology, rAAV
packaging nowadays does not rely on the presence of helper viruses such as
adenovirus or Herpes. Instead by using a helper-free system, rAAV can be
packaged easily by co-transfecting rAAV cis plasmid with Rep/Cap plasmid and
helper plasmid to a packaging cell. The helper plasmid will be simulating helper
virus by offering adenoviral E1, E2, E4, and VA RNAs during rAAV packaging. rAAV
is then produced in the packaging cell and can be harvested 2 to 3 days after
co-transfection via lysing packaging cells by 3xfreeze/thaw cycle. The rAAVs
released into supernatant is then purified via ultracentrifugation (Figure 5).
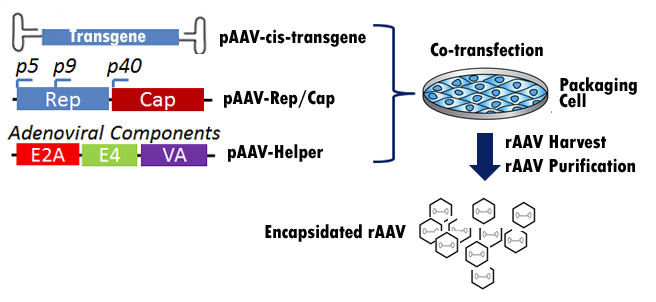
Figure 5.
A cartoon showing how rAAV is packaged with a helper free system.
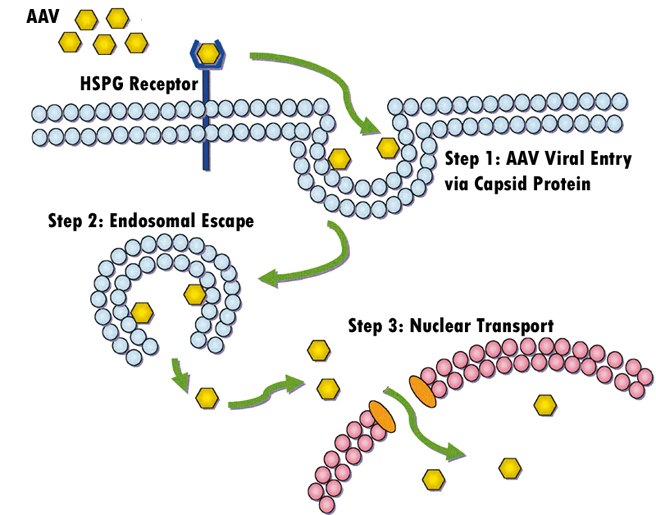
AAV Life Cycle:
Wild type AAV undergoes productive infection in
the presence of adenovirus co-infection. This is characterized by genome
replication, viral gene expression, and virion production. In the absence of
adenovirus, AAV can establish latency by integrating into chromosome 19 (AAVS1).
The latent AAV genome can be rescued and replicated upon superinfection by
adenovirus. Both stages of AAV's life cycle are regulated by complex
interactions between the AAV genome and AAV, adenoviral, and host proteins
(Figure 6).
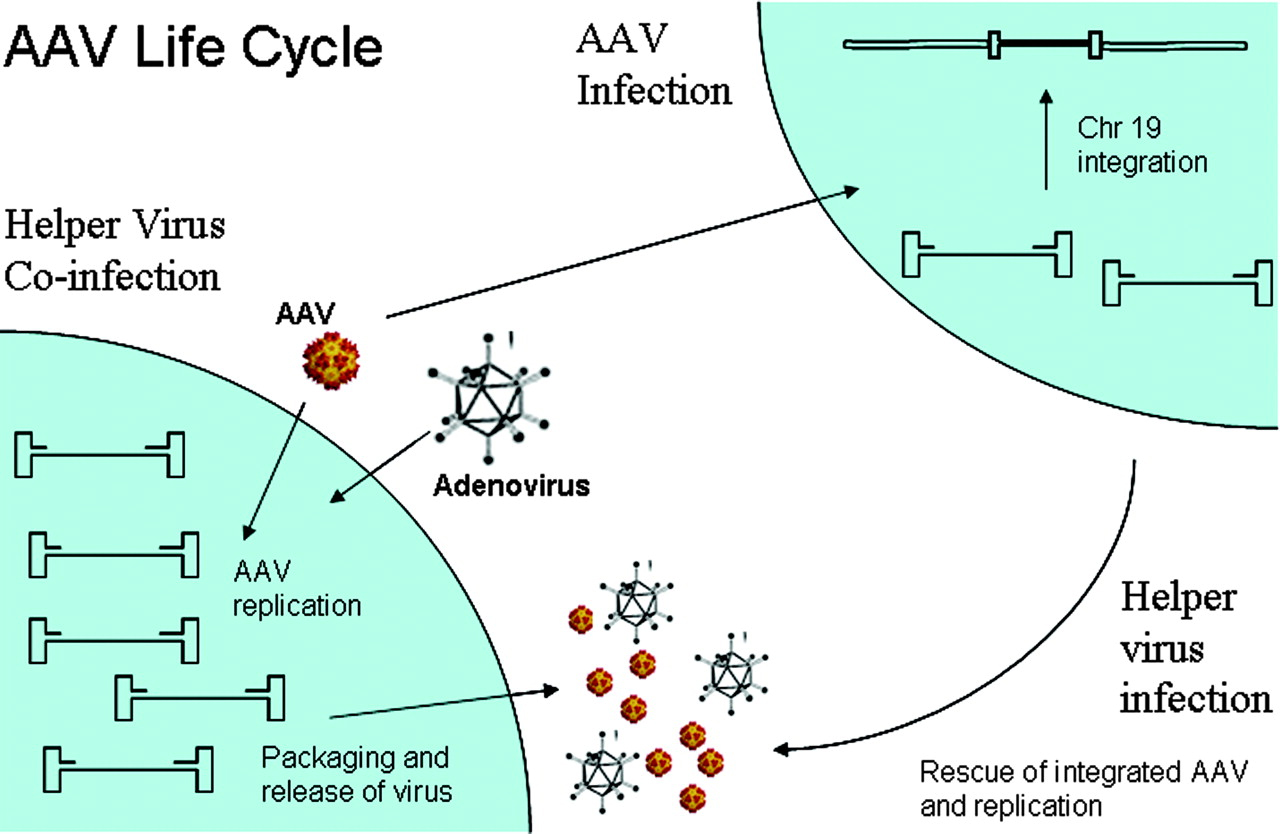
Figure 6.
A cartoon showing AAV life cycle.
Clinical Trials with rAAV Vectors:
To date, AAV vectors have been used for first- and second-phase clinical trials
for treatment of cystic fibrosis and first-phase trials for hemophilia.
Promising results have been obtained from phase I trials for Parkinson's
disease, showing good tolerance of an AAV2 vector in the central nervous system.
Other trials have begun, concerning AAV safety for treatment of Canavan disease,
muscular dystrophy and late infantile neuronal ceroid lipofuscinosis (table 2).
Table 2.
Selection of Clinical Trials using AAV-Based Vectors
|
Indication |
Gene |
Route of Administration |
Phase |
Subject Number |
Status |
|
Cystic fibrosis
|
CFTR |
Lung, via aerosol |
I |
12 |
Complete |
|
CFTR |
Lung, via aerosol |
II |
38 |
Complete |
|
|
CFTR |
Lung, via aerosol |
II |
100 |
Complete |
|
|
Hemophilia B
|
FIX |
Intramuscular |
I |
9 |
Complete |
|
FIX |
Hepatic artery |
I |
6 |
Ended |
|
|
Arthritis |
TNFR:Fc |
Intraarticular |
I |
1 |
Ongoing |
|
Hereditary emphysema |
AAT |
Intramuscular |
I |
12 |
Ongoing |
|
Muscular dystrophy |
Sarcoglycan |
Intramuscular |
I |
10 |
Ongoing |
|
Parkinson's |
GAD65, GAD67 |
Intracranial |
I |
12 |
Complete |
|
Canavan's |
AAC |
Intracranial |
I |
21 |
Ongoing |
|
Batten's |
CLN2 |
Intracranial |
I |
10 |
Ongoing |
|
Alzheimer's |
NGF |
Intracranial |
I |
6 |
Ongoing |
Trials for the treatment of prostate cancer
have reached phase III, however these ex vivo studies do not involve direct
administration of AAV to patients.
- Home
- :: Privacy Policy :: Terms & Conditions :: Contact Us
-
